Research Report
Cloning and Expression Analysis of IKKε Homologous Gene from Pinctada martensii 
 Author
Author  Correspondence author
Correspondence author
International Journal of Marine Science, 2014, Vol. 4, No. 56 doi: 10.5376/ijms.2014.04.0056
Received: 12 Jul., 2018 Accepted: 28 Jul., 2018 Published: 12 Oct., 2014
Jiao Y., Du X.D., Wang Q.H., Huang R.L., and Deng Y.W., 2014, Cloning and expression analysis of IKKε homologous gene from Pinctada martensii, International Journal of Marine Science, 4(56): 1-6 (doi: 10.5376/ijms.2014.04.0056)
IKKε (inhibitor of NF-κB kinases ε) is a key member in the NF-κB (nucleur factor κB) signal pathway and participates in the regulation of the cell differentiation, development, apoptosis and immune response. In this study, primers were designed based on the unigene sequence annotated as IKKε in the transcriptome database of Pinctada martensii, using rapid amplification of cDNA ends (RACE) technology, the full length sequence of IKKε gene was obtained from Pinctada martensii (PmIKKε). At the same time, the mRNA expression of PmIKKε in 6 tissues of Pinctada martensii was detected by Real-time PCR technology. The results showed that the obtained full length of PmIKKε gene cDNA was 2,843 bp, among which 5' UTR was 116 bp and 3' UTR was 516 bp, containing 27 bp polyA tail. The open reading frame (ORF) was 2,211 bp encoding 737 amino acid residues. The predicted molecular weight was 85.07 kD, isoelectric point was 6.17. Multiple sequence alignment indicated that PmIKKε had highly homology among other species IKKε and had 45% sequence identity with Crassostrea gigas. The software analysis results showed that the N-terminal region contained a protein kinase functional domain and an activation site of mitogen-activated protein kinase (MAPK), The C-terminal region contained a leucine zipper (LZ) domain and a helix-loop-helix (HLH) motif. Real-time PCR analysis demonstrated that PmIKKε was constitutively expressed in six studied tissues (hepatopancreas, gonad, hemolymph, gill, mantle, aductor muscle) in P. martensii, and the highest expression was found in hepatopancreas. This study could provide a theoretical basis for the further study of the function of PmIKKε in growth, development and innate immunity in of Pinctada martensii.
Background
Nuclear factor κB is a kind of nuclear transcriptional factor which widely exists in eukaryotic cells. It is the convergence point of various signal pathways and is involved in regulating cell proliferation, apoptosis, differentiation, stress and immune response (Zeng et al., 2007; Yang et al., 2011; Huang et al., 2012). In unstimulated cells, NF-κB binds to IκB (inhibitor of NF-κB) protein, which prevented NF-κB from entering the nucleus to play the role of transcription factor (Zhang et al., 2009; Yang et al., 2011). IκB kinase is a serine/threonine protein kinase family. At present, it was found that IKKs (IκB kinases, IKKs) consisted of five members: IKKα, IKKβ, and IKKε with catalytic activity, IKKγ, and IKAP (IKK complex-associated protein) with no catalytic activity. IKKs could phosphorylate specific sites on IκB, make IκB ubiquitin and eventually be degraded by proteasome, and remove the inhibitory effect of IκB on NF-κB, so that NF-κB entered the nucleus to play the role of transcription factor (Xiong et al., 2008). IKKε (IkB kinase epsilon) was one of the newly discovered members of IKK family. A serine/threonine kinase domain was found at the end of N-terminus, like α/β, earlier discovered member of the IKK family (Li, 2012). It had been found that IKKε could not only activate NF-κB by phosphorylation of IκB, but also directly bind to and phosphorylate IAP1 (inhibitor of apoptosis proteins 1, IAP1) when it was overexpressed to a certain extent in Drosophila melanogaster, which made IAP1 lose its ability to inhibit caspases. In an indirect way, the formation of normal cell morphology and other physiological processes were affected (Bergmann, 2006).
Pinctada martensii is the main shellfish for the cultivation of seawater pearls in China. Some scholars had analyzed the IκB (Zhang et al., 2009), NF-κB homologues (Wu et al., 2007; Huang et al., 2012) of Pinctada martensim, and detected their reaction under the stimulation of lipopolysaccharide LPS and Vibrio alginolyticus. Xiong et al. (2018) had found IKK homologues (Pf-IKK) in Pinctada martensii, and their sequence was 39% homologous with IKK- alpha of mice. It had 66% homology with the IKK-alpha of Crassostrea gigas. These studies indicated that there might be a typical NF-κB signaling pathway in Pinctada martensii. IKKε is one of the newly discovered members of the IKK family, which has not been reported yet. In this study, the full-length sequence of IKKε gene (PmIKKε) was obtained by RACE technology, and the expression of PmIKKε in six tissues of Pinctada martensii was detected by real-time PCR. The aim of this would provide a theoretical basis for further elucidation of the role of NF-κB signaling pathway in various physiological processes in the Pinctada martensii.
1 Results and Analysis
1.1 The cloning and sequence analysis of PmIKKε gene
According to the specific primers, which was the unigene sequence annotated IKKε in the transcriptome database of pearl sac of Pinctada martensii, this study adopted 5' RACE to amplified 5' upstream sequence of IKKε gene, and adopted 3'RACE to amplified 3' downstream sequence of IKKε gene. Meanwhile, this study obtained 186 bp 5' upstream sequence and 2,282 bp 3' downstream sequence respectively. The full-length cDNA sequence of 2,843 bp Pinctada martensii IKKε gene was obtained by splicing the sequence (Figure 1). This sequence 5' UTR was 116 bp, 3' UTR was 516 bp, and contained 27 bp polyA tail, the open reading frame was 2,211 BP, its molecular weight might be 85.07 kD, and the isoelectric point might be 6.17.
|
Figure 1 Sequence analysis of PmIKKε gene in Pinctada martensii Note: Capital letters represent coding sequence (ORF); Lowercase letters represent non-coding sequences (UTR); Protein kinase domain is shown in gray frame (13~283); MAPKK activation sites are underlined (237-241); Ltalics and underscores indicate the domain of leucine zipper; Helix-loop-helix (HLH) motif is shown in colorless frame (587~661) |

1.2 The amino acid sequence analysis of PmIKKε gene
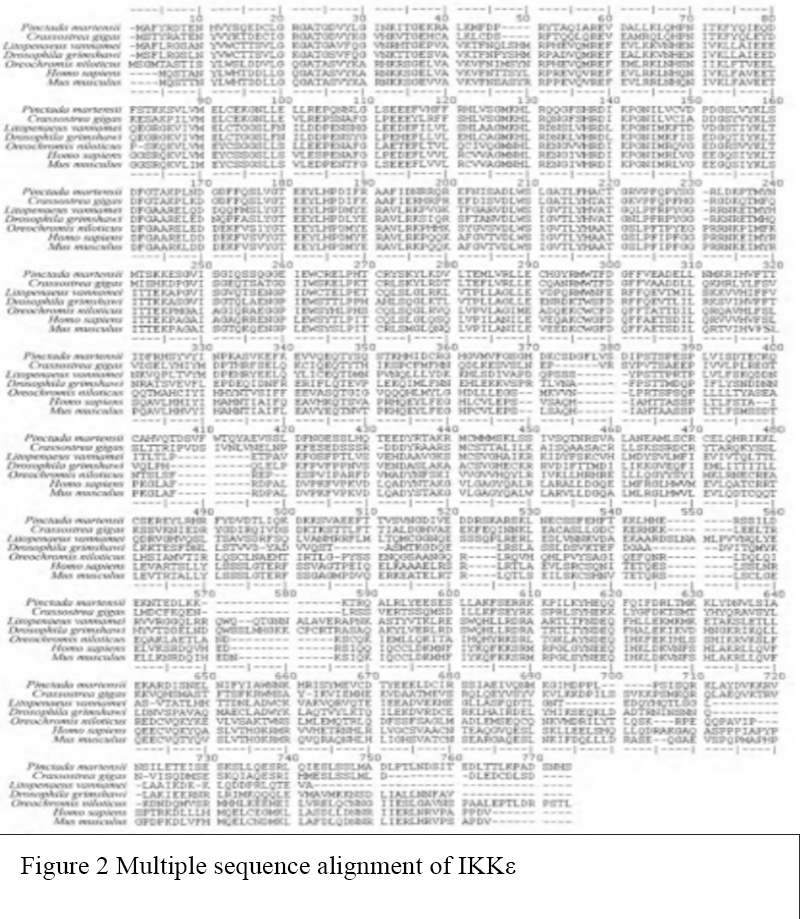
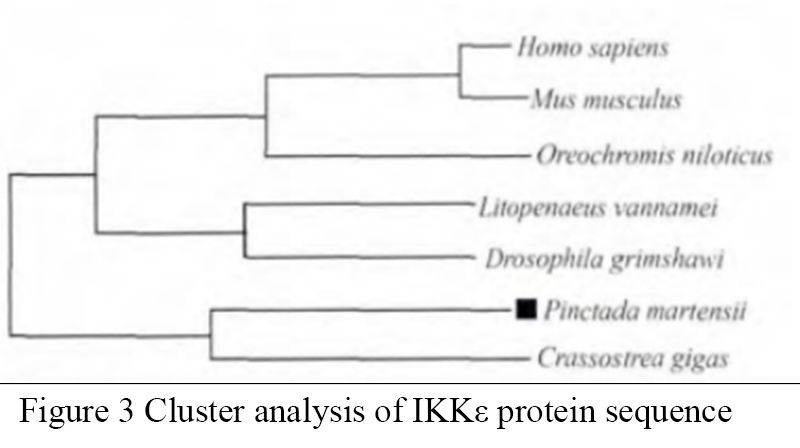
PmIKKε could encode 737 amino acids. The results of software analysis showed that the N-terminal of PmIKKε contained a functional domain of protein kinase and an activation site of MAPKK. There was a leucine zipper and a helix-loop-helix at the C-terminal (Figure 1). This study used clusterW5.0 to analyze the IKKε sequence of Pinctada martensii and the IKε protein sequence of other species. The results showed that the IKKε among different species had highly conservative, and the sequence similarity between PmIkeε and oyster was 45%. The conserved region was located in the protein kinase domain of the N-terminal of IKKε protein. The similarity of PmIKKε with the reported IKK homologue of Pinctada martensii is only 27% (Figure 2), which indicates that PmIKKε may belong to different IKK family with the reported IKK homologue. This study used MEGA5.1 software to analyze the LST8 protein sequences of Pinctada martensii, oyster, litopenaeus vannamei, drosophila, tilapia, mouse and human. As shown in Figure 3, the Pinctada martensii and oysters are clustered together, people, mouse and tilapia gather in one branch, drosophila and litopenaeus vannamei gather in one branch, and the clustering results are consistent with the traditional classification.
|
Figure 2 Multiple sequence alignment of IKKε |
|
Figure 3 Cluster analysis of IKKε protein sequence |
1.3 Quantitative tissue analysis of PmIKKε
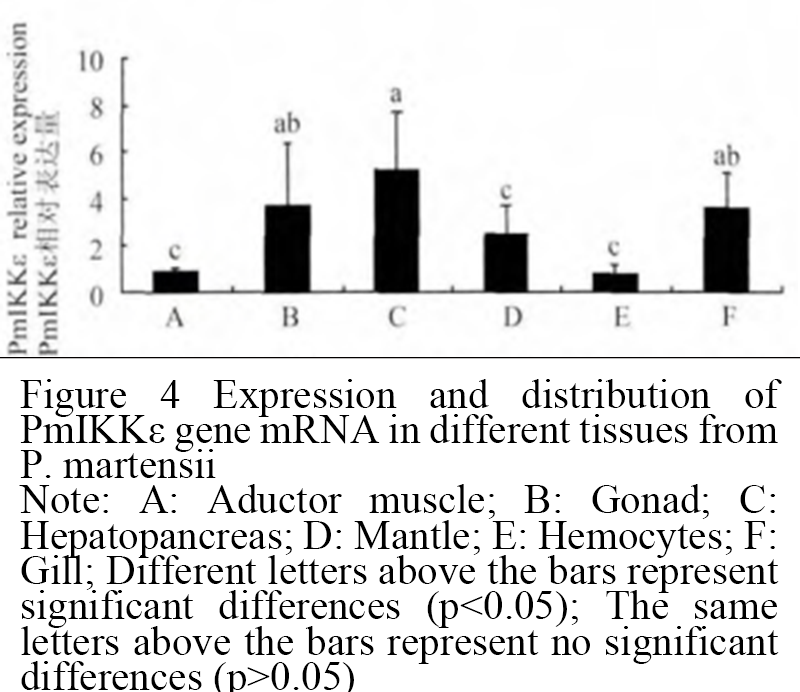
The expression of mRNA, which is the IKKε gene of Pinctada martensii in six tissues was detected by Real-time PCR. The results showed that PmIKKε was expressed in the coat membrane, hepatopancreas, gill, pearl sac, adductor muscle and gonad of Pinctada martensii. However, there was a significant difference in the expression level, and its expression in hepatopancreas was higher than that in the other tissues, then were gonads and gills (Figure 4).
|
Figure 4 Expression and distribution of PmIKKε gene mRNA in different tissues from P. martensii Note: A: Aductor muscle; B: Gonad; C: Hepatopancreas; D: Mantle; E: Hemocytes; F: Gill; Different letters above the bars represent significant differences (p<0.05); The same letters above the bars represent no significant differences (p>0.05) |
2 Discussion
The nuclear transcription factor NF-κB of eukaryotes widely existed in various biological cells from insect to human, and participates in the regulation of cell differentiation, development, apoptosis, adhesion and inflammation reaction. IKKs played an important role in the activation of NF-κB. When the cells were stimulated by external signals, activated IKKs phosphorylated IκB, and phosphorylated IκB was ubiquitized and degraded, thus causing NF-κB to remove the inhibitory effect of IκB. The IKKs family had five members: Ikeα, IKKβ, IKKε, IKKγ and IKAP. In mammals, Ikeε was similar to IKKα and IKKβ, and there was a kinase domain at the N terminal, which could phosphorylate IκB and activated NF-κB signaling pathway (Peters et al., 2000). The N-terminal of PmIKKε of Pinctada martensii obtained in this study also had a kinase domain. The multi-sequence alignment showed that the domain was highly conserved among species, which showed the importance of the domain. In addition, the C-terminal of the protein sequence of PmIKKε also contained a LZ and an HLH structure, just like the member structure of IKKs family. LZ was a structural unit that mediated the DNA-binding proteins between other proteins. The HLH structure was composed of a short α-helix through one ring and another long α-helix. HLH and LZ structures were the basis to form homodimer or heterodimer in the IKK family (Kwak et al., 2000). This information indicated that the sequence and structural characteristics of PmIKKε might have high homology with members of IKKs family.
The expression patterns of genes in different tissues would provide reference for the study of gene function. In order to further study the function of PmIKKε in Pinctada martensii, we analyzed the expression patterns of mRNA, which is the gene of PmIKKε in six different tissues of Pinctada martensii. The results showed that PmIKKε was expressed in six tissues of Pinctada martensii, with the highest expression in hepatopancreas. The results showed that PmIKKε might be involved in a wide range of functions in Pinctada martensii. Hepatopancreas might be shellfish immune organs that secrete various immune enzymes to counteract foreign interference. The function of hepatopancreas might be particularly important for shellfish without specific immunity (Tiscar and Mosca, 2004). It was reported that the NF-κB signaling pathway mediated by IKKε could induce the expression of purple phosphatase and superoxide dismutase (Darville et al., 2000; Matsumoto et al., 2001). We speculated that IKKε could activate NF-κB signaling pathway and promote hepatopancreas to secrete immune enzymes to counteract external stimuli. These results suggested that the hepatopancreas might have an active NF-κB signaling pathway compared with other tissues, which might further demonstrate the important role of hepatopancreas in the congenital immunity of shellfish.
To sum up, the full-length cDNA sequence of PmIKKε, a new member of the IKKε family, was obtained by race based on the transcriptome data of Eupatorium martensii and its function was preliminarily studied. Our results showed that PmIKKε might be very similar to the members of the IKK family at molecular level and function. This study might provide a basis for further study on the role of PmIKKε and its NF-κB signaling pathway in innate immunity of the Pinctada martensii.
3 Materials and Methods
3.1 Experimental material
The mantle membrane of Pinctada martensii was the material of RACE. The total RNA of 6 tissues from the adductor muscle, mantle membrane, gill, hepatopancreatic hemolymph and gonad of the adult Pinctada martensii were extracted as experimental materials of Real-time quantitative PCR, and at least 5 parallel individuals were taken from each tissue.
3.2 Strain and plasmid
Cloning vector pMD18-T Vector and competent JM109 were purchased from TaKaRa.
3.3 Main reagents
La Taq enzyme and RACE reagent box (clontech) were purchased from TaKaRa, M-MLV RTase cDNA Synthesis Kit and Trizol were purchased from Invitrogen, DyNAmoTM Color Flash SYBR Green qPCR kit was purchased from Thermo. The other reagents were domestic analytical pure reagent.
3.4 The full length sequence of race target gene
This study designed IKKε gene specific primers, according to the unigene sequence, which annotated IKKε in the transcriptome database of Pinctada martensii. The operations of 5' RACE and 3' RACE were designed according to the manual of SMARTTM race Amplification kit instruction of clontech. The obtained target fragment was purified and ligated to pMD18-T vector, then transformed into JM109 receptive cells. Positive clones were selected by colony PCR and sequenced. The full-length cDNA of the target gene was obtained by splicing the sequence with the known fragment (Jiao et al., 2012). The specific primers used are shown in Table 1.
|
Table 1 Primers for gene clone and fluorescence quantification of PmIKKε gene of Pinctada martensii Note: A: 5' RACE outer primer; B: 5' RACE inner prime; C: 3' RACE outer primer; D: 3' RACE inner primer; E: qPCR forward primer; F: qPCR reverse primer |
.png)
3.5 Bioinformatics analysis of target gene
Sequencing results and known unigene sequences were matched and spliced by bl2seq (http://www.ncbi.nlm.nih.gov/blast/bl2seq/wbl-ast2.cgi), and obtained the full-length cDNA sequences of each gene; this study predicted the coding frame and non-coding region by ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html), molecular weight and isoelectric point were analyzed on the Expasy website (http://web.expasy.org/compute_pi/).
3.6 The detection of the target gene expression by Real-time quantitative PCR
The expression of mRNA, which is the gene of IKKε in six tissues of Pinctada martensii was detected by Real-time quantitative PCR. Beta-actin was used as an internal reference gene. The results of Real-time quantitative PCR were analyzed by ABI step one software system. The reproducibility of samples and difference between groups were compared by SPSS 16.0.
Authors’ contributions
DXD was the principal of this project who directed the experimental design, completed and revised article. JY completed the experimental design, data analysis, finished article and revised it. WQ was responsible for gene clone and the experiment of Real-time quantitative PCR; HSL and DYW participated in the revision of article. All the authors read and agreed the final passage.
Acknowledgements
This study was funded by the National Natural Science Foundation of China (31272635, 412061-41, 31372526), the National Natural Science Foundation of Guangdong (S2012040008-042), the talent project of the department education in Guangdong (2012LYM_0074), and the PhD startup project in Guangdong Ocean University (1212318).
Bergmann A., 2006, IKK epsilon signaling: Not just NF-kappaB, Curr. Biol., 16(15): R588-R590
https://doi.org/10.1016/j.cub.2006.07.010
PMid:16890515
PMCid:PMC2601670
Darville M.I., Ho Y.S., and Eizirik D.L., 2000, NF-kappaB is required for cytokine-induced manganese superoxide dismutase expression in insulin-producing cells, Endocrinology, 141(1): 153-62
https://doi.org/10.1210/endo.141.1.7268
PMid:10614634
Huang X.D., Liu W.G., Guan Y.Y., Shi Y., Wang Q., Zhao M., Wu S.Z., and He M.X., 2012, Molecular cloning and characterization of class Ⅰ NF-kappaB transcription factor from pearl oyster (Pinctada fucata), Fish Shellfish Immunol., 33: 659-66
Jiao Y., Wang H., Du X., Zhao X., Wang Q., Huang R., and Deng Y., 2012, Dermatopontin, a shell matrix protein gene from pearl oyster Pinctada martensii, participates in nacre formation, Biochem Biophys Res, Commun., 425(3): 679-683
https://doi.org/10.1016/j.bbrc.2012.07.099
PMid:22842462
Kwak Y.T., Guo J., Shen J., and Gaynor R.B., 2000, Analysis of domains in the IKKalpha and IKKbetaproteins that regulate their kinase activity, J. Biol. Chem., 275(19): 14752-14759
https://doi.org/10.1074/jbc.M001039200
PMid:10747982
Li H.B., 2012, IKKε expression in human gliomas and its molecular mechanisms in promoting glioma cell proliferation and invasion, Thesis for M.S., Tianjin Medical University, Supervisor: Huang Q., pp.95-103
Matsumoto M., Hisatake K., Nogi Y., and Tsujimoto M., 2001, Regulation of receptor activator of NF-kappaB ligand-induced tartrate-resistant acid phosphatase gene expression by PU.1-interacting protein/interferon regulatory factor-4, Synergism with microphthalmia transcription factor, J. Biol. Chem., 276(35): 33086-33092
https://doi.org/10.1074/jbc.M104299200
PMid:11425859
Peters R.T., Liao S.M., and Maniatis T., 2000, IKK epsilon is part of a novel PMA-inducible IkappaB kinase complex, Mol. Cell, 5(3): 513-522
https://doi.org/10.1016/S1097-2765(00)80445-1
Tiscar P.G., and Mosca F., 2004, Defense mechanisms in farmed marine molluscs, Vet. Res. Commun., 28(Suppl. 1): 57-62
https://doi.org/10.1023/B:VERC.0000045379.78547.23
PMid:15372930
Wu X., Xiong X., Xie L., and Zhang R., 2007, Pf-Rel, a Rel/nuclear factor-kappaB homolog identified from the pearl oyster, Pinctada fucata, Acta Biochim. Biophys. Sin. (Shanghai), 39: 533-539
https://doi.org/10.1111/j.1745-7270.2007.00306.x
Xiong X., Feng Q., Chen L., Xie L., and Zhang R., 2008, Cloning and characterization of an IKK homologue from pearl oyster, Pinctada fucata, Dev. Comp. Immunol., 32(1): 15-25
https://doi.org/10.1016/j.dci.2007.03.013
PMid:17568671
Yang J.Y., Xu X.F., and Xiang Z.Y., 2011, Progress in research on NF-κB, Huaihai Yiyao (Journal of Huaihai Medicine), 29(1): 93-96
Zeng P., Deng H., and Liu L.M., 2007, New member of the IkB kinases: IKKε, Shijie Huaren Xiaohua Zazhi (World Chinese Journal of Digestology), 15(33): 3524-3526
Zhang D., Jiang S., Qiu L., Su T., Wu K., Li Y., Zhu C., and Xu X., 2009, Molecular characterization and expression analysis of the IkappaB gene from pearl oyster Pinctada fucata, Fish Shellfish Immunol., 26(1): 84-90
https://doi.org/10.1016/j.fsi.2008.10.009
PMid:19010421



. PDF(0KB)
. HTML
Associated material
. Readers' comments
Other articles by authors
. Xiaodong Du
. Yu Jiao
. Qingheng Wang
. Ronglian Huang
. Yuewen Deng
Related articles
. IKKε
. Pinctada martensii
. Gene clone
. Real-time PCR
Tools
. Email to a friend
. Post a comment